Биоэнергетика
В курсе “Биоэнергетика” рассматриваются основные сведения о механизмах преобразования энергии в живых организмах. Будут изложены системы субстратного, фото- и окислительного фосфорилирования, поставляющие всю энергию, необходимую для жизнедеятельности организмов. Основное внимание уделено процессам генерации мембранных форм энергии, а также их использования для синтеза ATФ, аккумуляции химических веществ в клетках и органеллах, подвижности бактерий, образования тепла в целях терморегуляции и т.д. В заключении будут рассмотрены пути практического применения биоэнергетики для отмены программы старения организма.
Курс предназначен для студентов IV курса. Курс лекций сопровождается проведением теоретических семинаров.
Лектор - академик РАН, д.б.н. В.П. Скулачев
Время проведения:Продолжительность курса:
Форма отчетности: Экзамен
Альтернативный курс:Нет
Программа курса
Вопросы к экзамену
Краткое введение в курс
Полное содержание курса изложено в учебном пособии “Мембранная биоэнергетика” В.П. Скулачев, А.В. Богачев, Ф.О. Каспаринский, Издательство Московского университета 2011 г.
Что связывает энергию и жизнь? В начале XX века Дж. Холдейн (Haldane) определил жизнь, как способ существования самовоспроизводящихся структур за счет притока энергии извне. Как следует из формулировки, способность к энергообеспечению является одним из двух основных свойств жизни. Однако каким образом энергия позволяет жизни существовать? Мы постараемся ответить на этот вопрос, по возможности не прибегая к помощи сложных формул.
Прежде всего, давайте определим тему нашего курса. Биоэнергетика (биологическая энергетика) есть совокупность процессов преобразования энергии внешних ресурсов в биологически полезную работу живых систем, а также раздел биологии, изучающий эти процессы. Истоками биоэнергетики можно считать рассуждения античных мыслителей о природе брожения и о роли воздуха в процессах использования пищи живыми организмами. Леонардо да Винчи одним из первых (начало XVI в.) сравнил питание животных с горением свечи. Эта идея была развита голландским естествоиспытателем Я.Б. Ван-Гелмонтом (1648 г.) в опытах с растениями. До начала ХХ века основным содержанием биоэнергетики являлось изучение суммарных балансов процессов энергообеспечения живых организмов (дыхания, брожения), а также воздействия различных условий (переход от покоя к работе, изменение окружающей температуры) на энергетический баланс организма. Быстрое развитие биохимии в первую половину ХХ века способствовало формированию представлений о превращениях энергии в живых клетках. Однако до формулирования П. Митчелом принципа устройства ключевых механизмов трансформации энергии биоэнергетика, как отдельная наука, еще не существовала.У каждой из современных отраслей науки есть свой год рождения. Так, молекулярная генетика ведет отсчет времени с публикации в «Nature» статьи Уотсона и Крика о двойной спирали. Биоэнергетика тоже имеет свою точку отсчета и тоже это статья в «Nature», но другого Нобелевского лауреата Питера Митчела. В 1961 г. он напечатал статью, где предложил объяснение механизма дыхательного и фотосинтетического фосфорилирования. А само слово «биоэнергетика» было предложено 5 годами раньше (1956 г.) Нобелевским лауреатом А. Сент-Дьерди, который дал такое название своей небольшой брошюре. Это слово, вынесенное на титульный лист, вошло в язык как название новой науки.
Выделившаяся в отдельную науку биоэнергетика быстро завоевала «место под солнцем». В 1978 г. получил Нобелевскую премию ее основатель П. Митчел. В 1988 г. Нобелевскими лауреатами стали Х. Михель, Й. Дейзенхофер и Р. Хубер, преуспевшие в рентгеноструктурном анализе фотосинтетических центров. В 1997 г П. Бойер и Дж. Уокер удостоились такой же премии за исследование протонной ATP-синтазы, а И.-Х. Скоу вместе с ними – за открытие Na+,K+-ATPазы. В 2003 г. Нобелевскими лауреатами стали П. Агре и Р. МакКаннон, описавшие структуру аквапорина и калиевого канала. Ждет своей премии открытие Г. Кремером, Д.Д. Ньюмейером и Кс. Уангом центральной роли митохондрий в запрограммированной смерти клетки. В последнее десятилетие число публикаций на эту тему растет по экспоненте.
Биоэнергетика в системе биологических наук
Классификацию биологических наук можно представить в виде здания, восемь этажей которого соответствуют уровням сложности организации живой материи. Самый верхний этаж – исследование биосферы (более высокие этажи пока только проектируются, поскольку неизвестно, есть ли жизнь за пределами нашей планеты). Спускаясь с этажа на этаж, исследователь продвигается от общего к частному. Второй этаж сверху – экология. Эта наука изучает сообщества отдельных организмов, которые образуют экосистемы, формирующие биосферу. Этажом ниже концентрируется группа наук, которым может быть объединено названием «биология видов». Это – классические описательные науки биологического профиля, такие как зоология, ботаника, микология, микробиология и вирусология. Они позволяют систематизировать представления о видах и популяциях на основании эталонных образцов или статистического анализа совокупности фенотипических признаков. Популяции состоят из индивидуумов. Их изучением занимаются на четвертом этаже, если считать сверху. Анатомия и физиология позволяют охарактеризовать соответственно строение и функции как целостного индивидуума, так и его тканей. Ткани состоят из клеток, и можно изучать биологические явления на этом субтканевом уровне (клеточная биология, 5-ый этаж сверху). Следующий уровень упрощения – субклеточная биология, объектами которой являются органеллы и надмолекулярные комплексы. Здесь объединены работы по исследованию структуры и функций плазмалеммы, цитозоля, клеточного ядра, митохондрий, эндоплазматического ретикулума, лизосом, пероксисом, хлоропластов и некоторых других внутриклеточных органелл.
Седьмой этаж сверху исключительно важен: это молекулярная биология. Предметом молекулярной биологии являются те индивидуальные молекулы, из которых состоят живые существа. Здесь прежде всего речь пойдет о макромолекулах. Именно на этом этаже происходит переход к количественному описанию биологических объектов. Биология молекул – хотя и очень сложная, но уже точная наука, претендующая на уровень точности, характерный для химии и физики. Непременным условием здесь является сохранение нативности исследуемых молекул и их комплексов: они должны быть «живыми», т.е. нативными, а не денатурированными, и продолжать во время эксперимента выполнение своей биологической функции. На самом нижнем этаже (8-ой сверху) это требование уже может не соблюдаться. Биоорганическая химия исследует структуру и физико-химические свойства чистых индивидуальных макромолекул, а также их компонентов и низкомолекулярных соединений, присутствующих в организме. Некоторые из таких соединений являются неорганическими, однако малочисленность объектов исследования не дает оснований для выделения бионеорганическох химии в отдельный этаж.
Классификация биологических наук не ограничивается уровнями сложности. Тот или иной биологический объект может исследовать биолог, химик, физик или математик. Каждый из них обладает совершенно разными методологическими подходами и философией исследования. Иногда даже сами выводы исследователя кардинальным образом зависят от того, в каком методологическом «подъезде здания биологии» он трудился. Для биоматематика важны точность измерений и их разброс. Биофизик, исследующий протекание реакций во времени, хочет наблюдать за ними с высоким разрешением на протяжении всего диапазона времен.
Для химика главное – чистота молекул и гомогенность системы. Биолог не будет в большинстве случаев стремиться к фемтосекундному разрешению, математической точности или химической чистоте. Биолога интересует поведение (регуляция, рождение и смерть) живой системы, а также ее история, имея в виду филогенез и онтогенез. Я по образованию – биолог и поэтому книга в значительной степени отражает биологический подход. Есть еще один критерий для классификации исследований в биологии, когда в основу положены те биологические функции, которые выполняют исследуемые системы. Существует четыре основные функции биологических объектов. Одна из них – генетическая, т.е. – передача свойств организма по наследству. Наука, изучающая эту функцию, называется генетикой. Энергообеспечение – другое необходимое условие существования живой системы, и этим занимается биоэнергетика. Есть такая функция, как превращение одних веществ в другие. Очевидно, что то, из чего мы состоим – это в основном совсем не те вещества, которые можно найти в окружающей среде. Большинство из них мы для себя сделали. Поэтому превращение веществ может быть расценено как специальная биологическая функция. С учетом соблюдения языковых норм ее можно назвать ее метаболикой. Это неологизм. В принципе, здесь речь идет о биохимии в ее прежнем понимании. Однако биохимия родила в свое время науки, занимающие в здании биологии три последних этажа: субклеточную биологию, молекулярную биологию и биоорганическую химию. В то же время, биохимию можно понимать как раздел биологии, в котором используется химический подход. И, наконец, сенсорика. Это тоже неологизм, который, по аналогии с тремя предыдущими, можно определить науку о восприятии и обработке сигналов, которые поступают из внутренней или из внешней среды живой системы, а также способность адекватно реагировать на такие сигналы.Следует подчеркнуть, что функциональный подход венчает собой изучение любой биологической системы. В конце концов, нам не так уж важно, какая конкретная молекула занимается каким-то делом, и уж совсем не важно, каким образом мы ее открыли – как биологи, химики, физики или математики. Важно, что именно делает молекула и как ее функция вписывается в общую схему жизнедеятельности исследуемого объекта. Эти четыре науки – цель биологии. Там, где мы решили задачу в ее функциональном аспекте, можно поставить точку и идти дальше.
Законы биоэнергетики
На сегодня биоэнергетика достигла такого прогресса в своем развитии, что мы уже можем сформулировать ее основные законы.
1-й закон биоэнергетики. Живая клетка избегает прямой утилизации энергии внешних ресурсов при совершении полезной работы. Сначала она трансформирует эту энергию в конвертируемую форму ATP, 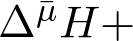 или
или 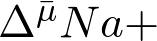 , которые и используются затем в различных энергоемких процессах (ур. 1). Апеллируя к нашей повседневной жизни, можно сказать, что клетка предпочитает бартеру денежное обращение.
, которые и используются затем в различных энергоемких процессах (ур. 1). Апеллируя к нашей повседневной жизни, можно сказать, что клетка предпочитает бартеру денежное обращение.
Энергетические ресурсы → ATP, или → работа (1)
Впервые этот принцип был выдвинут еще в 1941 г. Ф. Липманом, который тогда знал о существовании только одной биологической «валюты» – ATP. Действительно, ATP – абсолютно универсальный признак жизни: нет живой клетки, где бы не было ATP. Но оказалось, что существуют еще две биологических «валюты»: разности электрохимических потенциалов ионов H+ и Na+ ( и ).
может существовать в двух формах: 1) электрической и 2) химической (иначе концентрационной и осмотической). Первая из них – трансмембранная разность электрических потенциалов (ΔΨ), вторая – трансмембранная разность концентраций ионов водорода (ΔрН).
Если между двумя отсеками, разделенными мембраной, существует разность электрических потенциалов, образованная, например, электрической батареей, то энергетически выгодным оказывается перемещение ионов Н+ из отсека, заряженного положительно, в отрицательно заряженный отсек. Другой способ достичь той же цели – закислить содержимое левого отсека относительно правого. В этом случае протонный ток слева направо также будет идти «под гору», но движущей силой окажется уже не ΔΨ, а ΔрН.
Потенциальная энергия, накопленная в виде ΔΨ или ΔрН, может быть утилизирована полезным образом, если поместить в мембрану устройство, сопрягающее перенос протона «под гору» с совершением какой-либо полезной работы.
Энергия, накопленная в форме 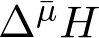 , может быть рассчитана по уравнению (2):
, может быть рассчитана по уравнению (2):
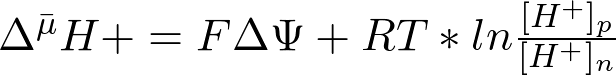 (2)
(2)
где ΔΨ – трансмембранная разность электрических потенциалов, R – газовая постоянная, T – абсолютная температура, F – число Фарадея, [H+]p и [H+]n – молярные концентрации ионов Н+ в соответственно положительно заряженном (или более кислом) и в отрицательно заряженном (или более щелочном) отсеках.
вычисляется в единицах джоуль х моль-1. Для перевода величины в вольты это значение необходимо разделить на число Фарадея. Полученное частное П. Митчел предложил называть протон-движущей силой (по аналогии с электродвижущей силой) и обозначил ее символом Δр. Величина Δр может быть рассчитана для 25оC по уравнению (3):
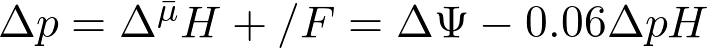 (3)
(3)
Согласно уравнению (3) ΔрН, равная 1, эквивалентна ΔΨ равной 0,06 В, или 60 мВ. Та же величина, выраженная в килоджоулях х моль-1, будет равна 5,7 (1,37 ккал х моль-1).
Аналогичные уравнения можно применить также и к натриевой энергетике. В этом случае в уравнение (3) вместо надо подставить , а вместо Δр – «натрий-движущую силу», которую можно обозначить как Δs (от лат. sodium):
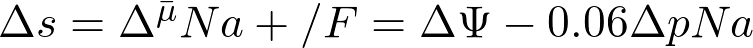 (4)
(4)
Свет или энергия субстратов дыхания могут утилизироваться ферментами фотосинтетической или дыхательной редокс-цепей или бактериородопсином. В результате образуется , которая затем используется для совершения работы, в частности для синтеза ATP. Субстратное фосфорилирование служит альтернативным механизмом образования ATP, который не требует . Такое фосфорилирование наблюдается в цепи реакций гликолиза и при окислительном декарбоксилировании α-кетоглутаровой кислоты.
-Зависимое образование ATP – главный, но не единственный процесс трансформации в химическую работу. К тому же типу энергетических превращений относятся синтез неорганического пирофосфата и перенос восстановительных эквивалентов в направлении более отрицательных редокс-потенциалов. Таковы обратный перенос электронов в дыхательной цепи и трансгидрогеназная реакция.
Зависящий от транспорт через мембрану различных форм веществ в сторону большей их концентрации представляет собой трансформацию энергии по типу → осмотическая работа, а вращение бактериального жгутика за счет энергии служит примером превращения → механическая работа. Образование теплоты митохондриями животных в ответ на понижение окружающей температуры описывается превращениями типа → теплопродукция.
Для немембранных частей клетки также описаны все перечисленные выше типы энергетических превращений. Здесь они поддерживаются энергией ATP или других высокоэнергетических соединений.
Существуют системы, специализированные на стабилизации (забуферивании) уровней и ATP. Для эту роль играют градиенты Na+ и K+, для ATP – креатинфосфат.
У некоторых бактерий вместо образуется . Источником энергии могут служить дыхание или неокислительное декарбоксилирование карбоновых кислот. Образованная используется для производства химической, осмотической и механической работы. Буфером может служить Н+/К+-градиент.
Наиболее сложный характер носит энергетика клетки животного. Фактически здесь в ходу три различные энергетические «валюты»: в митохондриях, на внешней клеточной мембране и ATP в прочих частях клетки.
Возвращаясь к 1-ому закону, нужно сказать, что три его компонента (ATP, , ) неравноправны. ATP – непременный компонент, а два остальных взаимозаменяемы и вследствие этого у некоторых организмов может присутствовать либо тот, либо другой компонент. Но один должен быть обязательно. Как следствие этого - 2-й закон биоэнергетики. Любая живая клетка располагает хотя бы двумя формами конвертируемой энергии – ATP и или .
3-й закон биоэнергетики предполагает взаимопревращение всех трех форм запасенной энергии. Вот почему клетка может удовлетворить все свои энергетические потребности, если есть возможность получить хотя бы одну из трех конвертируемых форм энергии за счет внешних энергетических ресурсов. Иными словами, клетке все равно, как расплачиваться за совершенную работу – банкнотами или кредитной карточкой.
Описаны клетки, использующие только какой-то один тип энергетических ресурсов. Одни из них живут за счет дыхания или фотосинтеза и образуют только , у них нет гликолиза, поэтому нет возможности прямого синтеза ATP. У других, наоборот, единственным источником энергии служит гликолиз, образующий ATP без посредничества . У некоторых бактерий нет ни гликолиза, ни дыхания, ни фотосинтеза: они накапливают за счет протекания одной из реакций декарбоксилирования, а из делают и ATP, и .
Мы обычно не осознаем масштаб событий, участвующих в энергообеспечении нашего организма. Вот некоторые цифры. Человек за день потребляет в среднем около 140 л кислорода и синтезирует около 40 кг ATP, для чего нужно перекачать через мембраны митохондрий порядка 0,5 кг протонов. При этом генераторы поддерживают напряженность электрического поля на митохондриальной мембране в сотни кВ/см.
В области науки, которой посвящен наш курс, еще есть много белых пятен. До сих пор не ясны молекулярные механизмы большинства взаимопревращений различных форм энергии. Ярчайшая иллюстрация глубины нашего незнания – история с открытием роли митохондрий в запрограммированной смерти клетки – апоптозе. В самом конце ушедшего ХХ века выяснилось, что в межмембранном пространстве митохондрий спрятаны «белки смерти», которые запускают апоптоз, выйдя из митохондрий в цитозоль. Таким образом, митохондрии не только электростанции клетки, но и органеллы, определяющие самую ее судьбу. А простейший из цитохромов – цитохром с, которым занимались, начиная с его открытия Д. Кейлиным в 1925 году, оказался мощным инструментом клеточного самоубийства, причем это совершенно другая функция данного белка, не имеющая отношения к его функции переносчика электронов.